внутрь бактериальной клетки (рисунок 32).
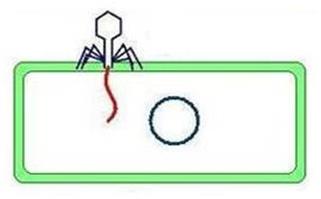
Рисунок 32 - Введение ДНК фага в бактериальную клетку.
После адсорбции происходит ферментативное расщепление клеточной стенки лизоцимом, находящимся в дистальной части фагового отростка. Одновременно в сократительном чехле бактериофага высвобождаются ионы кальция, которые активируют АТФазу. В результате этого происходит сокращение чехла и вталкивание стержня хвостового отростка через цитоплазматическую мембрану внутрь клетки. По каналу стержня ДНК фага впрыскивается в цитоплазму бактериальной клетки. Поскольку диаметр стержневого канала незначительно превышает диаметр молекулы ДНК (около 20 нм), то ДНК попадает в цитоплазму в линейной форме. При этом капсид и отросток остаются вне клетки.
Около 10% фаговой ДНК активно впрыскивается внутрь клетки во время сокращения чехла, а остальная часть ДНК втягивается в цитоплазму бактерии благодаря процессам транскрипции и трансляции.
При адсорбции фагов на половых ворсинках проникновение нуклеиновой кислоты происходит по каналу F-пили.
2. Биосинтез компонентов фага внутри бактериальной клетки (рисунок 34).

Рисунок 34 - Биосинтез компонентов фага.
Инъецированная нуклеиновая кислота вызывает перестройку метаболизма бактериальной клетки: прекращается синтез бактериальных нуклеиновых кислот и белков. В эту стадию ДНК бактериофага транскрибируется с помощью собственной транскриптазы и образуется иРНК, которая поступает на рибосомы клетки-хозяина, где синтезируются фаговые белки.
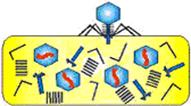
3. Сборка структурных компонентов фаговых частиц (рисунок 35).
Рисунок 35 – Сборка структурных компонентов фаговых частиц.
Вновь синтезированные белки в цитоплазме клетки формируют капсиды, отростки и другие структуры дочерних фаговых частиц. После этого синтезированная нуклеиновая кислота фагов заполняет пустотелые капсиды головок.
4.
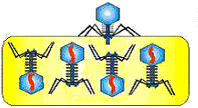
Сборка (морфогенез) зрелых фаговых частиц (рисунок 36).
Рисунок 36 - Сборка зрелых фаговых частиц.
Заполненный нуклеиновой кислотой капсид взаимодействует с хвостовой частью и образует зрелую фаговую частицу. Соединение хвоста с головкой происходит только после полной их сборки. Точно так же ворсинки присоединяются к хвосту только после полного соединения отростка с головкой. Благодаря строгому генетическому контролю со стороны фага обеспечивается последовательность и согласованность всех процессов его внутриклеточного размножения. После сборки в каждой бактериальной клетке накапливается до 200 фаговых частиц (рисунок 37).
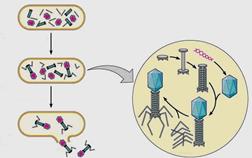 Рисунок 37 – Сборка фаговых частиц
Рисунок 37 – Сборка фаговых частиц
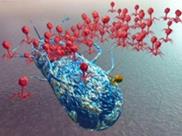
1. Выход фаговых частиц из клетки (рисунок 38).
Рисунок 38 - Выход фаговых частиц из клетки.
Вновь образованные фаговые частицы выходят наружу в результате лизиса клеточной оболочки изнутри фаговым лизоцимом. Он синтезируется на последнем этапе размножения фага. Некоторые нитевидные фаги высвобождаются из клетки путем прохождения нуклеиновой кислоты через цитоплазматическую мембрану и клеточную стенку бактерий. Во время такого прохождения фаги приобретают капсиды. Один цикл от момента адсорбции фага на поверхности клетки до выхода потомства из клетки продолжается 30-40 минут. Такие циклы продолжаются до тех пор, пока не будут лизированы все чувствительные к фагу клетки.
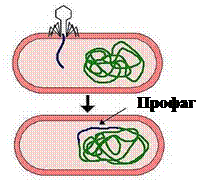
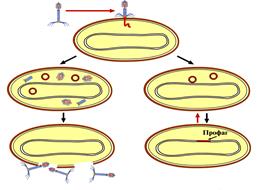
ДНК умеренных фагов после проникновения в цитоплазму встраивается в геном бактериальной клетки. ДНК фага, встроенная в хромосому бактерии, называется
профагом, а культура таких бактерий –
лизогенной культурой (рисунок 39).
Рисунок 39 - Лизогенная культура бактерий.
Совместное сосуществование бактерии и умеренного бактериофага называется лизогенией (от греч. lysis - разложение, genea - происхождение). Разли- чают монолизогенность и полилизогенность. Бактериальная культура, образующая после индукции один вид фага, называется монолизогенной, а бактериальная культура, образующая несколько видов фагов, называется полилизогенной.
ДНК умеренного фага встраивается в строго определенную область генома, синхронно с ним реплицируется и передается по наследству. При лизогении фагового потомства не образуется, так как в бактерии синтезируется специфический низкомолекулярный белок репрессор, подавляющий транскрипцию фаговых генов. Биосинтез репрессора детерминируется генами профага. Лизогенные бактерии приобретают иммунитет (невосприимчивость) к последующему проникновению и размножению в них родственного фага.
У некоторых лизогенных культур спонтанно или под действием физических факторов и химических веществ (УФ-лучи, ионизирующее излучение, перекисные соединения, митомицин С и др.) профаг может исключаться из хромосомы и становиться литическим фагом. Этот процесс называется индукцией профага. Он заканчивается продукцией фагов и лизисом бактерий.
Лизогенизация бактерий может приводить к изменению их морфологических, культуральных, ферментативных, антигенных и биологических свойств. При этом профаг может придавать бактерии новые ранее отсутствовавшие у нее свойства. Феномен изменения свойств микробов под влиянием профага называется фаговой конверсии (от лат. conversio - превращение). Например, профаг придает дифтерийной палочке способность продуцировать экзотоксин.
Таким образом, различают следующие типы взаимодействия фагов с бактериями:
- продуктивный тип взаимодействия, при котором в клетке образуется фаговое потомство, обусловливающее лизис бактерий;
- интегративный тип взаимодействия, при котором нуклеиновая кислота фага встраивается в геном бактерии и сосуществует с ним в течение длительного времени.
Схема продуктивного и интегративного типов взаимодействия бактериофага и клетки представлена на рисунке 40.
Лизис Лизогения
Рисунок 40 - Схема продуктивного и интегративного путей развития фага.
Некоторые умеренные фаги могут быть дефектными, то есть неспособными образовывать зрелые фаговые частицы ни в естественных условиях, ни при индукции профага.