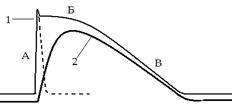
 Потенциал действия (1) и кривая сокращения (2) гладкомышечной клетки.
Потенциал действия (1) и кривая сокращения (2) гладкомышечной клетки.
А – фаза деполяризации (Na+- вход);
Б – «кальциевое плато» (Ca2+- вход);
В – фаза реполяризации (К+- выход); (пунктиром ПД скел. мышцы)
Особенности сокращения. Процесс сокращения гладкомышечных волокон совершается по тому же механизму скольжения нитей актина и миозина относительно друг друга, что и в скелетных мышцах, однако у гладкомышечных клеток нет той стройной оранжировки сократительных белков, как у скелетных мышц. У этих клеток миофибриллы с саркомерами расположены нерегулярно, поэтому клетка не имеет поперечной исчерченности. Электромеханическое сопряжение в гладкомышечных клетках идет иначе, чем в скелетных мышцах, так как в них слабо выражен саркоплазматический ретикулум. В связи с этим триггером для мышечного сокращения служит поступление ионов Са2+ в клетку из межклеточной среды в процессе генерации ПД. Того количества кальция, которое входит в клетку при возбуждении, вполне достаточно для полноценного фазного сокращения.
Инициация сокращений гладких мышц с помощью ионов Са2+ также имеет несколько иной механизм, чем в п-п. волокнах. Ионы Са2+ воздействуют на белок кальмодулин, который активирует киназы легких цепей миозина. Это обеспечивает перенос фосфатной группы на миозин и сразу вызывает сокращение поперечных мостиков. О существовании тропонин-тропомиозиновой системы сведений нет. При снижении в миоплазме концентрации ионов Са2+ фосфатаза дефосфорилирует миозин и он перестает связываться с актином. Скорость сокращения гладких мышц невелика, она на 1 — 2 порядка ниже, чем у скелетных мышц. Сила сокращений некоторых гладких мышц позвоночных не уступает силе сокращений скелетных мышц.
 Механизм возбуждения гладкой мышцы. ПП некоторых гладкмыш. волокон, обладающих автоматией, обнаруживает постоянные небольшие колебания, обеспечивающие постоянные небольшие колебания, обеспечивающие возникновение ПД при достижении КП. Величина мембранного потенциала меньше значения мембранного потенциала меньше значения мембранного потенциала скелетных мышц и составляет в волокнах, не обладающих автоматией, - -60…-70 мВ, она несколько ниже в спонтанно активных клетках - -30…-70 мВ.
Механизм возбуждения гладкой мышцы. ПП некоторых гладкмыш. волокон, обладающих автоматией, обнаруживает постоянные небольшие колебания, обеспечивающие постоянные небольшие колебания, обеспечивающие возникновение ПД при достижении КП. Величина мембранного потенциала меньше значения мембранного потенциала меньше значения мембранного потенциала скелетных мышц и составляет в волокнах, не обладающих автоматией, - -60…-70 мВ, она несколько ниже в спонтанно активных клетках - -30…-70 мВ.
ПД. В гладких мышцах внутренних органов регистируются ПД 2х основных типов: пикоподобные ПД и ПД с выраженным плато. Длительность пикоподобных плато ПД составляет 5-80 мс, ПД с плато, характерное для гладких мышц мптки, уретры и некоторых сосудов, длятся 30-500 мс.
Ионный механизм возникновения ПД в гладких мышцах: при удалении из омывающего раствора ионов Na+ (замена их ионами Li+ или холина) возникают полноценные ПД у гладких мышц. Удаление из раствора ионов Са2+ или воздействие на мышечные клетки блокаторов кальциевых каналов (например, верапамила) ведет к обратимому угнетению ПД. Все эти факты свидетельствуют о главной роли ионов Са2+ в генерации ПД гладких мышц. Электровозбудимые медленные Са-каналы обладают меньшей ионной избирательностью, чем «быстрые» натриевые каналы нервных и поперечно-исчерченных мышечных волокон. Помимо двухвалентных катионов они проницаемы и для ионов Na+.
ПД гладких мышц, состоящие из начального пикового компонента и последующего плато, имеют более сложную ионную природу. Например, в гладких мышцах мочеточников начальный фрагмент ПД имеет преимущественно кальциевую природу, а последующий медленный компонент (плато) — преимущественно натриевую природу.







